Biologia 5: luku 7 Mutaatiot
Mutaatioita syntyy sekä DNA:n kopiointivirheinä että ympäristötekijöiden vaikutuksesta
Mutaatiot
ovat perimässä tapahtuvia pysyviä muutoksia. Mutaatiot ovat elämän jatkumisen
edellytys! Ilman niitä ei evoluutiota ja luonnon monimuotoisuutta olisi. Mutaatioita
voi tapahtua yksittäisessä geenissä, kromosomissa tai koko kromosomistossa. Jos
DNA:n korjaamisesta huolehtivat entsyymit eivät kykene paikkaamaan
vaurioitunutta osaa, muutos jää pysyväksi. Tällöin sitä kutsutaan mutaatioksi.
Mutageenit
ovat mutaatioita aiheuttavia ympäristötekijöitä. Esim ionisoiva säteily
(röntgen- ja gammasäteily), UV-säteily, monet kemikaalit sekä eräät virukset. Lisäksi
luonnossa syntyvät tai synteettisesti valmistetut kemialliset yhdisteet
aiheuttavat mutaatioita. Esim bentseeni ja kadmiumyhdisteet.
Lyhytaaltoinen
säteily voi aiheuttaa mutaation hajottamalla vesimolekyylin vetyioniksi ja
hydroksyyliradikaaliksi. Hydroksyyliradikaali korvaa nopeasti menettämänsä ionin
ottamalla sen DNA-juosteesta. Tupakan savu sisältää paljon mutageenejä.

Albiinotiikeri.
Kuva Googlen kuvahaku.
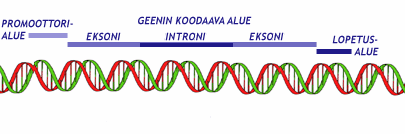
Geenimutaatiossa yksittäisen geenin rakenne muuttuu
Geenimutaatio
tapahtuu yhdessä geenissä. Sen seurauksena geenin sisältämä perinnöllinen
informaatio muuttuu, minkä seurauksena voi olla muutos proteiinin rakenteessa. Pienin
mahdollinen muutos geenissä on pistemutaatio eli yhden nukleotidin muuttuminen
toiseksi. Yleensä pistemutaatio on harmiton. Jos pistemutaatio esiintyy
yleisenä populaatiossa, puhutaan yhden emäksen vaihtelusta eli snipistä. Geenimutaatioissa
voi nukleotidejä myös hävitä tai tulla ylimääräisiä. Pistemutaatio ei aina
muuta proteiinin aminohappokoostumusta.
Jos
pistemutaation seurauksena jokin proteiinin rakenteen kannalta keskeinen
aminohappo muuttuu toiseksi, voivat solun rakenteeseen ja toimintaan
kohdistuvat seuraukset olla huomattavia. Esim sirppisoluanemiassa yhden
aminohapon muuttuminen toiseksi aiheuttaa koko erytrosyytin muodon muuttumisen
kiekkomaisesta sirppimäiseksi, jolloin erytrosyytin hapenkuljetuskyky
heikkenee.
Eräät
hyvin vakavat sairaudet johtuvat geenissä olevien nukleotidien häviämisestä.
Esim lähes yksinomaan pojilla esiintyvä perinnöllinen lihasrappeumatauti
(Duchennen lihasdystrofia) johtuu sukusolun X-kromosomissa tapahtuneesta
geenimutaatiosta. Tässä mutaatiossa lihassolujen tukiproteiinia tuottavasta
geenistä on hävinnyt nukleotidejä, mikä aiheuttaa ennenaikaisen lopetuskolmikon
syntymisen lähetti-RNA:han ja estää toimivan proteiinin synnyn.
Kromosomimutaatiossa kromosomin rakenne muuttuu
Kromosomimutaatiossa
kokonainen kromosomin osa muuttuu. Seurauksena on muutos geenien määrässä tai
rakenteessa. Kromosomimutaatioita on viittä tyyppiä, ja ne ovat häviämä,
liittymä, siirtymä, kääntymä ja kahdentuma. Häviämässä kromosomista irtoaa
pala, eikä sitä liitetä takaisin irtoamispaikkaan. Häviämät vahingoittavat
lähes poikkeuksetta solun toimintaa.
Liittymässä
kromosomista irronnut pala siirtyy saman kromosomin eri paikkaan tai kokonaan
toiseen kromosomiin. Siirtymässä kahdesta eri kromosomista on samanaikaisesti
irronnut pala, ja palat vaihtavat paikkoja keskenään. Suurimmalla osalla näistä
kromosomimutaatioista ei kuitenkaan ole vaikutusta soluihin.
Kääntymässä
kromosomista irtoaa pala, joka kääntyy korjauksen yhteydessä toisin päin.
Yleensä kääntymällä ei ole merkitystä alueen geenien toimintaan. Kahdentumassa
jokin kromosomin osa monistuu. Myös transposonit eli hyppivät geenit
aiheuttavat kromosomimutaatioita. Siirtyessään jonkin geenin sisään tai sen
säätelyalueelle transposonit aiheuttavat muutoksia tämän geenin toimintaan. Transposonien
arvellaan olevan peräisin viruksista. Sukusoluissa esiintyvät
kromosomimutaatiot voivat aiheuttaa sairauksia jälkeläiselle. Usein nämä
mutaatiot aiheuttavat epämuodostumia ja johtavat keskenmenoon.
Kromosomistomutaatiossa kromosomiluku muuttuu
Kromosomistomutaatiossa
joko yksittäisten kromosomien tai koko kromosomiston lukumäärä muuttuu.
Kromosomistomutaatio johtuu yleensä meioosissa tapahtuneesta häiriöstä. Kasveilla
tavataan varsin yleisesti yksilöitä, joiden kromosomiluku on muuttunut
lajiristeymän seurauksena. Monosomiassa toinen jonkin kromosomiparin
vastinkromosomeista puuttuu. Vastaavasti mutaatiota, jossa
vastinkromosomipareja on kahden sijasta kolme, kutsutaan trisomiaksi.
Turnerin
oireyhtymässä toinen X-kromosomi puuttuu. Tästä on seurauksena mm. sukupuolisen
kehityksen viivästyminen. Jo yksi ylimääräinen tai puuttuva kromosomi on
yleensä sitä kantavalle yksilölle vahingollinen. Tällöin myös alleeleita on
poikkeava määrä, mikä vaikuttaa geenin ilmentymiseen.
Polyploidiassa
koko peruskromosomisto monistuu. Monisoluiset eliöt ovat yleensä diploidisia
(2n) eli niillä on kaksi peruskromosomistoa. Polyploidiassa
peruskromosomeja on enemmän.
Triploidiassa niitä on kolme (3n) ja tetraploidiassa neljä (4n) jne. Kasveilla
polyploidia on yleistä, koska niillä ylimääräinen kromosomisto ei yleensä
haittaa yksilönkehitystä, toisin kuin esim eläimillä.
Polyploidiaa
on kahdenlaista, autopolyploidia ja allopolyploidia. Autopolyploidiassa lajin
peruskromosomisto on monistunut, jolloin yksilössä on jokaisesta geenistä
enemmän kuin kaksi alleelia. Allopolyploidiassa kromosomistot ovat peräisin
kahdesta keskenään risteytyneestä lajista.
Mutaatiot vaikuttavat evoluutioon
Periytyvät
mutaatiot eli sukusolulinja soluissa tapahtuvat mutaatiot aiheuttavat yhdessä
rekombinaation kanssa populaation perinnöllisen muuntelun, joka ilmenee yksilöiden
erilaisissa rakenteissa, elintoiminnoissa ja käyttäytymisessä. Parhaiten
ympäristöönsä sopeutuneet yksilöt kykenevät tuottamaan eniten
lisääntymiskykyisiä jälkeläisiä, ja niiden geenit alkavat yleistyä
populaatiossa.
Uusi
alleeli syntyy, kun geenin rakenne muuttuu. Mitä vähemmän yksilöitä
populaatioon kuuluu, sitä suuremmalla todennäköisyydellä jälkeläinen perii
kaksi kyseistä alleelia, ja vasta tällöin vaikutus näkyy yksilön fenotyypissä.
Kromosomistomutaatioilla on suuri merkitys kasvien evoluutiossa
Polyploidia
lisää kasvien kykyä sopeutua vaativiin ympäristöolosuhteisiin, koska
moninkertainen geenistö takaa nopean kasvun esim silloin, kun kasvukausi on
lämpötilan tai veden saannin vuoksi lyhyt. Ylimääräinen tai puuttuva kromosomi
haittaa enemmän solun toimintaa kuin kaikkien kromosomien lukumäärän muutos. Allopolyploidia
voi tuottaa uuden lajin yhdellä hyppäyksellä ilman maantieteellistä
isolaatiota.
Eläimillä
polyploidia on harvinaista, koska ylimääräinen kromosomisto aiheuttaa vakavia
kehityshäiriöitä yksilönkehityksen aikana ja johtaa esim ihmisellä
keskenmenoon. Hedelmöittymättömästä munasolusta lisääntymistä kutsutaan
partenogeneesiksi.